The Dons' Organic Garden
- Thread starter DonPetro
- Start date



DonTesla
Well-Known Member
Cheers bro, looking forward! Almost there..im def gunna do that cheers once my tab stops neing a pain I am going 2 take a pic of one of my deep dreams this one is in a small pot in coco but dam shes really starting 2 sparkle now I think im around week 6 now




DonTesla
Well-Known Member

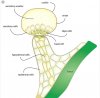
Note the expression here.. Various hues of pink, purple, magenta and black "Secretory cells"..
This time we had no negative pressure and no VP drops.. And as predicted ..no hypodermal cells affected.
Also no surprise that the basal cell (see below) also remained unaffected, as usual, thus far anyway
However, the interesting thing is that We were able to achieve this much expression with our water only soil recipe .. and the real kicker, no uv!!
.
DonTesla
Well-Known Member
Cyanidin, delphinidin, pelargonidin, peonidin, malvidin, and petunidin are the most common anthocyanidins distributed in the plants. The distribution of these anthocyanidins in fruits and vegetables is 50%, 12%, 12%, 12%, 7%, and 7%, respectively [3]. Their molecular structures are shown in Figure 3. In nature, cyanidin is a reddish-purple (magenta) pigment. It is the major pigment in berries [4] and other red-colored vegetables such as red sweet potato and purple corn [5]. Delphinidin has a chemical characteristic similar to most of the anthocyanidins. It appears as a blue-reddish or purple pigment in the plant. The blue hue of flowers is due to the delphinidin pigment [6]. Pelargonidin differs from most of the anthocyanidins. In nature, it appears as red-colored pigment [7]. Pelargonidin gives an orange hue to flowers [8] and red to some of the fruits and berries [9].
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5613902/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5613902/
DonTesla
Well-Known Member


Antioxidants
The health and therapeutic effects of anthocyanin are mainly contributed by its antioxidative activities. As reported in the literature [105], anthocyanin chalcones and quinoidal bases with a double bond conjugated to the keto group are efficient antioxidants in scavenging free radicals. Also, the glycosylated B-ring structure of anthocyanin contributes to the high antioxidant activity, where ortho-hydroxylation and methoxylation substantially increase the antioxidant activity [106].
In fact, anthocyanidin has a higher ORAC value than anthocyanin. One of the possible reasons is anthocyanin aglycone is very unstable and highly reactive [107]. Anthocyanin, with the addition of an extra sugar at position C-3 in the heterocyclic C-ring, has lower antioxidant activity than the anthocyanidin with a single sugar molecule [108]. Acylation of anthocyanin with phenolic acid has a significant increase in antioxidant activity [109]. Diacylation of the anthocyanin markedly increases the antioxidant activity but 5-glycosylation leads to a reduction in the activity [107].
A previous study reports the antioxidant activity of malvidin-3-glucoside that was determined by metal-catalyzed lipid peroxidation models in comparison with other antioxidants [30]. The result shows that the quinoidal-base and pseudo-base of malvidin-3-glucoside significantly inhibited peroxidation of linoleate by myoglobin compared with catechin. In the presence of hydrogen peroxide-activated myoglobin, malvidin-3-glucoside had the highest antioxidant activity, followed by catechin, malvidin, and resveratrol. In term of glycosylated anthocyanin, addition of an extra glucose to cyanidin-3-xylosyl-galactoside forms cyanidin-3-xylosyl-glucosyl-galactoside with an ORAC value lower than the anthocyanin without addition of an extra sugar [107]. Acylation of malvidin-3-glucoside with p-coumaric acid has antioxidant activity assessed by linoleic acid oxidation higher than the non-acylated counterpart [109].
Anthocyanins have many other therapeutic effects in addition to their antioxidant activities. As an active pharmaceutical ingredient, anthocyanin pigment, such as delphinidin, has been patented for several therapeutic effects. Delphinidin is well-known for combating melanoma cells [110], as well as antimicrobial effects, such as curing Staphylococcus aureus infection [111]. It has also been used as the source of antiphlogistic or immunosuppressive active ingredients [112].
Literature shows that anthocyanins extracted from plants have antioxidative properties. Pelargonidin-3-glucoside, cyanidin-3-glucoside, and delphinidin-3-glucoside isolated from Phaseolus vulgaris L. (black bean) seed coat, as well as their standard aglycones, have strong antioxidative activity in a liposomal system and reduced formation of malondialdehyde by UVB irradiation [113]. The study also indicates that delphinidin and delphinidin-3-glucoside had the highest inhibitory effect on lipid peroxidation and O2•− scavenging activity. On the contrary, pelargonidin had the highest inhibitory effect on hydroxyl radical scavenging activity.
On the other hand, a study demonstrates that cyanidin and cyanidin-3-glucoside have the highest inhibitory effect on copper (II)-induced low-density lipoprotein (LDL) oxidation compared with the other phenolic acids, anthocyanins, and anthocyanin aglycones, whereas delphinidin has intermediate efficacy [114]. Comparing the result from both studies, the second study does not determine the efficacy of pelargonidin to inhibit lipid peroxidation.
DonTesla
Well-Known Member
Angiogenesis and development of diseases
Endothelial cells are the main cells involved in the angiogenesis process. Disturbances in physiologic angiogenesis can contribute to various human diseases, including CVDs, cancer, and diabetic complications such as diabetic retinopathy and nephropathy [115]. Normal angiogenesis depends on the intricate balance between angiogenic (VEGF, FGF2-fibroblast growth factor, TGF-β-transforming growth factor, and angiopoietin) and antiangiogenic (angiostatin, endostatin, and thrombospondins) factors [116].
The antiangiogenic effect of anthocyanins has been reported by several studies. Anthocyanin-rich extracts of several berries (wild blueberry, bilberry, cranberry, elderberry, and strawberry) significantly suppress hydrogen peroxide and TNF-α-induced vascular endothelial growth factor (VEGF) expression in HaCaT cells (human keratinocytes) [117]. Bilberry anthocyanidins (delphinidin, cyanidin, and malvidin) are also reported to inhibit VEGF-induced tube formation in a co-culture of human umbilical vein endothelial cells and fibroblasts [118].
Anthocyanin-rich purple corn extract attenuates endothelial expression of VEGF and hypoxia inducible factor (HIF)-1α, as well as to induce endothelial marker of platelet endothelial cell adhesion molecule-1 and integrin β3 induced by high glucose condition in human renal mesangial and endothelial cells [87]. Also, glomerular angiogenesis in the diabetic kidneys of db/db mice is disrupted by weakening the induction of VEGF and HIF-1α in vivo. The purple corn extract also diminishes the mesangial and endothelial induction of angiopoietin proteins under hyperglycemic conditions. These findings suggest that anthocyanin-rich purple corn extract antagonizes glomerular angiogenesis in high blood glucose condition through disturbing the Angpt-Tie-2 ligand-receptor system linked to the renal VEGF receptor-2 signaling pathway.
Cardiovascular health
Epidemiological studies show the relationships between anthocyanin-rich foods and CVDs, as well as the relationship between total anthocyanin intake and risk of developing these cardiovascular-related diseases. Anthocyanins also demonstrate in vitro anti-thrombotic effect [71]. The anti-thrombotic effect is supported by another study that anthocyanin-containing maize seed (20% seed in the diet) fed rats for eight weeks are less susceptible to ischemia-reperfusion injury and reduction of infarct size with increased myocardial antioxidant enzyme [73]. Also, Bell and Gochenaur [72] reveal that anthocyanin-rich extracts of chokeberry and bilberry, but not elderberry, possess vasorelaxation properties. Moreover, there is also no alteration of coronary response to nitric oxide which is a potent vasodilator agent.
In a clinical trial, the researchers suggest that consumption of anthocyanin-rich strawberries for one month improves lipid profile and platelet function in healthy volunteers [74]. Nonetheless, the effects may be attributed to the presence of non-anthocyanin compounds in strawberries, such as vitamin C and phenolic compounds. Moreover, the study should have control groups for comparison. However, Curtis et al. [119] indicate the consumption of 500 mg/day of elderberry extract for 12 weeks is ineffective in reducing the risk of CVD in healthy postmenopausal women. There is also no change in metabolic processing following 12 weeks of elderberry intake compared with acute intake [120].
Endothelial cells are the main cells involved in the angiogenesis process. Disturbances in physiologic angiogenesis can contribute to various human diseases, including CVDs, cancer, and diabetic complications such as diabetic retinopathy and nephropathy [115]. Normal angiogenesis depends on the intricate balance between angiogenic (VEGF, FGF2-fibroblast growth factor, TGF-β-transforming growth factor, and angiopoietin) and antiangiogenic (angiostatin, endostatin, and thrombospondins) factors [116].
The antiangiogenic effect of anthocyanins has been reported by several studies. Anthocyanin-rich extracts of several berries (wild blueberry, bilberry, cranberry, elderberry, and strawberry) significantly suppress hydrogen peroxide and TNF-α-induced vascular endothelial growth factor (VEGF) expression in HaCaT cells (human keratinocytes) [117]. Bilberry anthocyanidins (delphinidin, cyanidin, and malvidin) are also reported to inhibit VEGF-induced tube formation in a co-culture of human umbilical vein endothelial cells and fibroblasts [118].
Anthocyanin-rich purple corn extract attenuates endothelial expression of VEGF and hypoxia inducible factor (HIF)-1α, as well as to induce endothelial marker of platelet endothelial cell adhesion molecule-1 and integrin β3 induced by high glucose condition in human renal mesangial and endothelial cells [87]. Also, glomerular angiogenesis in the diabetic kidneys of db/db mice is disrupted by weakening the induction of VEGF and HIF-1α in vivo. The purple corn extract also diminishes the mesangial and endothelial induction of angiopoietin proteins under hyperglycemic conditions. These findings suggest that anthocyanin-rich purple corn extract antagonizes glomerular angiogenesis in high blood glucose condition through disturbing the Angpt-Tie-2 ligand-receptor system linked to the renal VEGF receptor-2 signaling pathway.
Cardiovascular health
Epidemiological studies show the relationships between anthocyanin-rich foods and CVDs, as well as the relationship between total anthocyanin intake and risk of developing these cardiovascular-related diseases. Anthocyanins also demonstrate in vitro anti-thrombotic effect [71]. The anti-thrombotic effect is supported by another study that anthocyanin-containing maize seed (20% seed in the diet) fed rats for eight weeks are less susceptible to ischemia-reperfusion injury and reduction of infarct size with increased myocardial antioxidant enzyme [73]. Also, Bell and Gochenaur [72] reveal that anthocyanin-rich extracts of chokeberry and bilberry, but not elderberry, possess vasorelaxation properties. Moreover, there is also no alteration of coronary response to nitric oxide which is a potent vasodilator agent.
In a clinical trial, the researchers suggest that consumption of anthocyanin-rich strawberries for one month improves lipid profile and platelet function in healthy volunteers [74]. Nonetheless, the effects may be attributed to the presence of non-anthocyanin compounds in strawberries, such as vitamin C and phenolic compounds. Moreover, the study should have control groups for comparison. However, Curtis et al. [119] indicate the consumption of 500 mg/day of elderberry extract for 12 weeks is ineffective in reducing the risk of CVD in healthy postmenopausal women. There is also no change in metabolic processing following 12 weeks of elderberry intake compared with acute intake [120].
DonTesla
Well-Known Member
Anticancer
Anthocyanins have been extensively studied for their anticancer properties, as well as antiangiogenesis, based on in vitro and cell culture studies, and animal models. Angiogenesis is the key for cancer development, where it is an important step in the transition of tumors from a benign state to a malignant one. In cancer prevention, antiangiogenesis is the process that prevents formation of new blood vessels that supply oxygen to the tumor cells. Several phytochemicals, including flavonoids and anthocyanins, are potential antiangiogenic agents.
Anthocyanins have been extracted and isolated from different plant sources for investigating their anticancer ability on esophagus, colon, breast, liver, hematological, and prostate cancers. The evidence from a previous study shows that 5% whole freeze-dried black raspberries and the anthocyanin-rich fraction supplemented to N-nitrosomethylbenzylamine-induced F344 rats have chemopreventive potential, where the treatment groups inhibit cell proliferation, inflammation, angiogenesis, and induce apoptosis in both preneoplastic and papillomatous esophageal tissues [75]. Thus anthocyanins have chemoprophylaxis potential.
Blueberry anthocyanins and anthocyanin-pyruvic acid adduct extracts (250 μg/ml) demonstrated anti-invasive potential in both breast cancer cell lines, MDA-MB-231 and MCF7 [76]. The extracts inhibited proliferation of cancer cell by acting as chemoinhibitors. The anthocyanin-pyruvic acid adduct extract has a better effect in MDA-MB-231, suggesting an effect independent of estrogen receptors. In addition to blueberry anthocyanins, anthocyanin-rich extracts (50 µg monomeric anthocyanin/ml) from chokeberry result in 60% growth inhibition of human HT-29 colon cancer cells within 24 h exposure, increase expression of tumor suppression genes (p21WAF1 and p27KIP1) and a 35% decrease in the cyclooxygenase-2 gene expression. As expected, the extracts have no obvious growth inhibition on normal colonic cell.
In another study, supplementation of anthocyanin-rich extracts of bilberry, chokeberry, and grape (containing 3.85 g anthocyanins per kg diet) for 14 weeks significantly reduced azoxymethane-induced aberrant crypt foci by 26–29% in 3–4 week-old male-specific pathogen-free F344 rats [80]. This reduction is associated with reduced cell proliferation and decreased expression of the COX-2 gene. The result also shows that the urinary 8-OHdG levels were similar among rats fed with different diets.
Dietary anthocyanin-enriched purple-fleshed sweet potato clone P40 significantly suppresses formation of aberrant crypt foci in the colons of female CF-1 mice coincided with a greater expression of apoptotic caspase-3 in the colon mucosal epithelial cells [81]. The observation suggests that anthocyanin-enriched sweet potato P40 has a protective effect against colorectal cancer by inducing cell-cycle arrest, anti-proliferative, and through apoptotic mechanisms. Another study also proves that anthocyanin extract (2 and 5 mg/ml) of purple potato induces maturation of acute myeloid leukemia cells via TNF-related apoptosis-inducing ligand [84]. Moreover, the less common anthocyanin source from vine was reported having anti-invasive property in human hepatoma Hep3B cells in a cancer study [83].
Similarly, anthocyanin-enriched black rice extract has an anticancer effect on breast cancer cells. The extract inhibited growth of breast cancer cell lines MCF-7 (ER+, HER2/neu−), MDA-MB-231 (ER−, HER2/neu−), and MDA-MB-453 (ER−, HER2/neu+) and induces apoptosis in MDA-MB-453 cells by depolarizing mitochondrial membrane potential and releasing cytochrome C into the cytosol, and thus triggered programmed cell death through apoptosis [77]. Oral administration of the same extract (100 mg/kg/day) to BALB/c nude mice bearing MDA-MB-453 cell xenografts significantly reduced tumor growth and suppresses angiogenesis by lowering the expression of angiogenesis factors matrix metallopeptidase-9, matrix metallopeptidase-2, and urokinase plasminogen activator in the tumor tissue. The results from both in vitro and in vivo studies suggest that anthocyanin-enriched black rice extract exhibits anticancer capability against human breast cancer cells by inducing cell apoptosis and suppressing angiogenesis.
In another study, black rice anthocyanins suppress metastasis in breast cancer cells by targeting the mitogen-activated protein kinase pathway [78]. The anthocyanins inhibited migration and invasion of MDA-MB-453 cells (HER2+), suppressed activation of rapidly accelerated fibrosarcoma, mitogen-activated protein kinase (MEK), and c-Jun N-terminal kinase (JNK), as well as downregulated secretion of matrix metalloproteinase 2 (MMP2) and MMP9. The study suggests that black rice anthocyanins suppress metastasis in breast cancer cells by targeting the RAS/RAF/MAPK (retrovirus-associated DNA sequences/rapidly accelerated fibrosarcoma/mitogen-activated protein kinase) pathway. Thus, it may be useful to treat patients at an advanced cancer stage.
Anthocyanins have been extensively studied for their anticancer properties, as well as antiangiogenesis, based on in vitro and cell culture studies, and animal models. Angiogenesis is the key for cancer development, where it is an important step in the transition of tumors from a benign state to a malignant one. In cancer prevention, antiangiogenesis is the process that prevents formation of new blood vessels that supply oxygen to the tumor cells. Several phytochemicals, including flavonoids and anthocyanins, are potential antiangiogenic agents.
Anthocyanins have been extracted and isolated from different plant sources for investigating their anticancer ability on esophagus, colon, breast, liver, hematological, and prostate cancers. The evidence from a previous study shows that 5% whole freeze-dried black raspberries and the anthocyanin-rich fraction supplemented to N-nitrosomethylbenzylamine-induced F344 rats have chemopreventive potential, where the treatment groups inhibit cell proliferation, inflammation, angiogenesis, and induce apoptosis in both preneoplastic and papillomatous esophageal tissues [75]. Thus anthocyanins have chemoprophylaxis potential.
Blueberry anthocyanins and anthocyanin-pyruvic acid adduct extracts (250 μg/ml) demonstrated anti-invasive potential in both breast cancer cell lines, MDA-MB-231 and MCF7 [76]. The extracts inhibited proliferation of cancer cell by acting as chemoinhibitors. The anthocyanin-pyruvic acid adduct extract has a better effect in MDA-MB-231, suggesting an effect independent of estrogen receptors. In addition to blueberry anthocyanins, anthocyanin-rich extracts (50 µg monomeric anthocyanin/ml) from chokeberry result in 60% growth inhibition of human HT-29 colon cancer cells within 24 h exposure, increase expression of tumor suppression genes (p21WAF1 and p27KIP1) and a 35% decrease in the cyclooxygenase-2 gene expression. As expected, the extracts have no obvious growth inhibition on normal colonic cell.
In another study, supplementation of anthocyanin-rich extracts of bilberry, chokeberry, and grape (containing 3.85 g anthocyanins per kg diet) for 14 weeks significantly reduced azoxymethane-induced aberrant crypt foci by 26–29% in 3–4 week-old male-specific pathogen-free F344 rats [80]. This reduction is associated with reduced cell proliferation and decreased expression of the COX-2 gene. The result also shows that the urinary 8-OHdG levels were similar among rats fed with different diets.
Dietary anthocyanin-enriched purple-fleshed sweet potato clone P40 significantly suppresses formation of aberrant crypt foci in the colons of female CF-1 mice coincided with a greater expression of apoptotic caspase-3 in the colon mucosal epithelial cells [81]. The observation suggests that anthocyanin-enriched sweet potato P40 has a protective effect against colorectal cancer by inducing cell-cycle arrest, anti-proliferative, and through apoptotic mechanisms. Another study also proves that anthocyanin extract (2 and 5 mg/ml) of purple potato induces maturation of acute myeloid leukemia cells via TNF-related apoptosis-inducing ligand [84]. Moreover, the less common anthocyanin source from vine was reported having anti-invasive property in human hepatoma Hep3B cells in a cancer study [83].
Similarly, anthocyanin-enriched black rice extract has an anticancer effect on breast cancer cells. The extract inhibited growth of breast cancer cell lines MCF-7 (ER+, HER2/neu−), MDA-MB-231 (ER−, HER2/neu−), and MDA-MB-453 (ER−, HER2/neu+) and induces apoptosis in MDA-MB-453 cells by depolarizing mitochondrial membrane potential and releasing cytochrome C into the cytosol, and thus triggered programmed cell death through apoptosis [77]. Oral administration of the same extract (100 mg/kg/day) to BALB/c nude mice bearing MDA-MB-453 cell xenografts significantly reduced tumor growth and suppresses angiogenesis by lowering the expression of angiogenesis factors matrix metallopeptidase-9, matrix metallopeptidase-2, and urokinase plasminogen activator in the tumor tissue. The results from both in vitro and in vivo studies suggest that anthocyanin-enriched black rice extract exhibits anticancer capability against human breast cancer cells by inducing cell apoptosis and suppressing angiogenesis.
In another study, black rice anthocyanins suppress metastasis in breast cancer cells by targeting the mitogen-activated protein kinase pathway [78]. The anthocyanins inhibited migration and invasion of MDA-MB-453 cells (HER2+), suppressed activation of rapidly accelerated fibrosarcoma, mitogen-activated protein kinase (MEK), and c-Jun N-terminal kinase (JNK), as well as downregulated secretion of matrix metalloproteinase 2 (MMP2) and MMP9. The study suggests that black rice anthocyanins suppress metastasis in breast cancer cells by targeting the RAS/RAF/MAPK (retrovirus-associated DNA sequences/rapidly accelerated fibrosarcoma/mitogen-activated protein kinase) pathway. Thus, it may be useful to treat patients at an advanced cancer stage.
DonTesla
Well-Known Member
Antidiabetes
The antidiabetic effect of anthocyanins from plants has been widely studied. Anthocyanin-rich Cornus fruits have been used in traditional Chinese prescription medicines to treat diabetes [121]. Primary bioactive components reported in Cornus fruits are the glycosides of cyanidin, delphinidin, and pelargonidin [98]. Jayaprakasam et al. [98] report that cyanidin-3-glucoside and delphinidin-3-glucoside effectively aided insulin secretion from rodent pancreatic β-cells (INS-1 832/13) in vitro compared with the other anthocyanins and anthocyanidins studied.
Another study demonstrates that pelargonidin and pelargonidin-3-galactoside caused a 1.4-fold increase in insulin secretion at 4 mM glucose concentration representative of the normal glucose level in human [122]. The ability of the anthocyanins to induce insulin secretion is in the increasing order of pelargonidin-3-galactoside, cyanidin-3-glucoside, and delphinidin-3-glucoside. This finding demonstrates that the number of hydroxyl groups on the B-ring of anthocyanins plays a crucial role in their ability to secrete insulin. Nevertheless, cyanidin, delphinidin, pelargonidin, malvidin, and petunidin do not potentiate significant insulin secretion.
In a clinical trial of 24 weeks involving 58 diabetic patients, the subjects in the anthocyanin group consumed two anthocyanin capsules (160 mg anthocyanins) twice daily purified from bilberry and blackcurrant [86]. The results show that anthocyanin group had a significantly lower fasting plasma glucose and insulin resistance index, as well as significantly elevated serum adiponectin and β-hydroxybutyrate concentrations compared to the placebo supplementation. However, the authors did not elucidate the mechanism involved for the prevention of insulin resistance in the diabetic patients.
Bilberry anthocyanin has been reported to ameliorate hyperglycemia and insulin sensitivity via activation of adenosine monophosphate-activated protein kinase (AMPK) in type 2 diabetic mice at skeletal muscle, liver, and white adipose tissue [85]. The activation of AMPK causes upregulation of glucose transporter 4 in the skeletal muscle and white adipose tissue while inhibiting glucose production in the liver. AMPK activation in the liver also results in a significant reduction in liver and serum lipid content via the phosphorylation of acetyl-CoA carboxylase (ACC), upregulation of peroxisome proliferator-activated receptor alpha (PPARα), acyl-CoA oxidase, and carnitine palmitoyltransferase-1A gene expressions.
It has been reported that a reduction in AMPK activity leads to diabetic nephropathy, which is associated with increased oxidative stress and lipid accumulation. Supplementation of anthocyanin-rich Seoritae extract restores AMPK activity, activates target molecules such as ACC, sterol regulatory element-binding protein 1, and PPAR, and suppresses intrarenal lipid accumulation in kidney tissue [88]. However, the authors did not examine specific contributions of the bioactive compounds in the Seoritae extract to the observed effects and the amounts of these compounds incorporated into the kidney. It is unsure if the target of anthocyanins is only AMPK or adiponectin. During the onsets of diabetic microangiopathic, microvascular permeability and the number of leucocytes sticking to the venular endothelium are increased [123]. In db/db mice, cyanidin-3-glucoside (2 g/kg diet) increases adiponectin secretion from adipose tissue, thus it protects the mice against diabetes-related endothelial dysfunction [89]. The study also shows that cyanidin-3-glucoside supplementation for eight weeks resulted in a noticeable improvement in endothelium-dependent relaxation of aorta of the mice.
The antidiabetic effect of anthocyanins from plants has been widely studied. Anthocyanin-rich Cornus fruits have been used in traditional Chinese prescription medicines to treat diabetes [121]. Primary bioactive components reported in Cornus fruits are the glycosides of cyanidin, delphinidin, and pelargonidin [98]. Jayaprakasam et al. [98] report that cyanidin-3-glucoside and delphinidin-3-glucoside effectively aided insulin secretion from rodent pancreatic β-cells (INS-1 832/13) in vitro compared with the other anthocyanins and anthocyanidins studied.
Another study demonstrates that pelargonidin and pelargonidin-3-galactoside caused a 1.4-fold increase in insulin secretion at 4 mM glucose concentration representative of the normal glucose level in human [122]. The ability of the anthocyanins to induce insulin secretion is in the increasing order of pelargonidin-3-galactoside, cyanidin-3-glucoside, and delphinidin-3-glucoside. This finding demonstrates that the number of hydroxyl groups on the B-ring of anthocyanins plays a crucial role in their ability to secrete insulin. Nevertheless, cyanidin, delphinidin, pelargonidin, malvidin, and petunidin do not potentiate significant insulin secretion.
In a clinical trial of 24 weeks involving 58 diabetic patients, the subjects in the anthocyanin group consumed two anthocyanin capsules (160 mg anthocyanins) twice daily purified from bilberry and blackcurrant [86]. The results show that anthocyanin group had a significantly lower fasting plasma glucose and insulin resistance index, as well as significantly elevated serum adiponectin and β-hydroxybutyrate concentrations compared to the placebo supplementation. However, the authors did not elucidate the mechanism involved for the prevention of insulin resistance in the diabetic patients.
Bilberry anthocyanin has been reported to ameliorate hyperglycemia and insulin sensitivity via activation of adenosine monophosphate-activated protein kinase (AMPK) in type 2 diabetic mice at skeletal muscle, liver, and white adipose tissue [85]. The activation of AMPK causes upregulation of glucose transporter 4 in the skeletal muscle and white adipose tissue while inhibiting glucose production in the liver. AMPK activation in the liver also results in a significant reduction in liver and serum lipid content via the phosphorylation of acetyl-CoA carboxylase (ACC), upregulation of peroxisome proliferator-activated receptor alpha (PPARα), acyl-CoA oxidase, and carnitine palmitoyltransferase-1A gene expressions.
It has been reported that a reduction in AMPK activity leads to diabetic nephropathy, which is associated with increased oxidative stress and lipid accumulation. Supplementation of anthocyanin-rich Seoritae extract restores AMPK activity, activates target molecules such as ACC, sterol regulatory element-binding protein 1, and PPAR, and suppresses intrarenal lipid accumulation in kidney tissue [88]. However, the authors did not examine specific contributions of the bioactive compounds in the Seoritae extract to the observed effects and the amounts of these compounds incorporated into the kidney. It is unsure if the target of anthocyanins is only AMPK or adiponectin. During the onsets of diabetic microangiopathic, microvascular permeability and the number of leucocytes sticking to the venular endothelium are increased [123]. In db/db mice, cyanidin-3-glucoside (2 g/kg diet) increases adiponectin secretion from adipose tissue, thus it protects the mice against diabetes-related endothelial dysfunction [89]. The study also shows that cyanidin-3-glucoside supplementation for eight weeks resulted in a noticeable improvement in endothelium-dependent relaxation of aorta of the mice.
DonTesla
Well-Known Member
Visual health
Anthocyanin pigments are important nutraceuticals in maintaining good vision. Anthocyanin-rich berries are traditionally known for the goodness to eyes and are often associated with night vision. Most of the berries have high anthocyanins content. Oral administration of bilberry extract (contained about 39% anthocyanins) to six weeks old C57BL/6 mice has been shown to prevent impairment of photoreceptor cell function during retinal inflammation [91]. In another study, 132 patients with normal tension glaucoma were supplemented with two anthocyanins capsules (60.0 mg anthocyanin in each capsule) from bilberry daily and have improved visual function, based on the Humphrey visual field test and minimal angle of resolution best-corrected visual acuity assessment [90].
Some other berries demonstrate a protective effect for eyesight. Blackcurrant anthocyanin supplementation (50 mg/day) for 24 months increased ocular blood flow in 19 patients with open-angle glaucoma, however, there were no significant changes in the intraocular pressure [95]. Supplementation of anthocyanins (50 mg/kg body weight) from the seed coat of black soybean to N-methyl-N-nitrosourea-induced retinal degenerative rats also prevents retinal degeneration [94], and also suppresses human lens epithelial cell death under hydrogen peroxide-induced oxidative stress by 50–200 μg/ml of the extract [93]. Anthocyanin also predominates around 70% in purple corn seed [124], where purple corn seed extract decreases lens opacity together with lower malonaldehyde levels [92].
Anti-obesity effect
Anthocyanidin and anthocyanin pigments possess anti-obesity properties. Based on a previous study, obese mice fed a diet rich in cyanidin-3-glucoside from purple corn for 12 weeks have reduced body weight, as well as decreases in white and brown adipose tissue weights [100]. The study demonstrates that hyperglycemia, hyperinsulinemia, hyperleptinemia, and an increase in the tumor necrosis factor (TNF-a) mRNA level occurred in the obese rats are normalized when treated with purple corn diet. The purple corn also suppresses mRNA levels of enzymes involved in fatty acid and triacylglycerol synthesis and lowered sterol regulatory element binding protein-1 mRNA level in the white adipose tissue. These downregulations may contribute to a low triacylglycerol accumulation in white adipose tissue.
Obesity is strongly associated with adipocyte dysfunction. Therefore, regulation of protein secretion from adipocyte or the adipocyte-specific gene expression is one of the most important targets for prevention of obesity. Tsuda and his research team further investigated the potency of anthocyanins, particularly cyanidin and cyanidin-3-glucoside on isolated rat adipocytes for anti-obesity effect. He demonstrates that the adipocytes treated with anthocyanins have increased adiponectin and leptin secretions and upregulated adipocyte-specific gene expression without activation of PPARγ in the isolated rat adipocytes [99]. Gene expression of adiponectin is also upregulated in white adipose tissue of the anthocyanin-treated mice. The increased phosphorylation of AMPK may be associated with these changes, and the monophosphate/adenosine triphosphate ratio is significantly decreased by the administration of anthocyanins.
As previously reported by Tsuda et al., adipocyte gene expression is not thoroughly studied. A further examination of gene expression profile in isolated rat adipocytes treated with anthocyanins (100 nM cyanidin-3-glucoside or cyanidin) has been performed in vitro [125]. Within 24 h, a total of 633 genes and 427 genes were upregulated (1.5-fold) by the treatment of adipocytes with cyanidin-3-glucoside and cyanidin, respectively. The upregulated genes include lipid metabolism and signal transduction-related genes. However, the altered genes are partially different when comparing the cyanidin-3-glucoside and cyanidin treated groups. They also report that treatment of adipocytes with cyanidin-3-glucoside and cyanidin upregulated hormone sensitive lipase and enhanced lipolytic activity based on the microarray data. Even though the findings have identified new responsive genes with potentially important functions in adipocytes related to obesity, additional investigation is needed. In vivo adipocytes are not likely to be exposed to the anthocyanidin due to its instability in the culture.
Another study found that the ameliorated obese mice (C57BL/6) fed with Cornelian cherries (Cornus mas) containing anthocyanins (1 g/kg of high fat diet) for eight weeks had a 24% decrease in weight gain and decreased lipid accumulation in the liver, as well as a significant decrease in liver triacylglycerol concentration, independent of food intake [126]. The diet containing a mixture of delphinidin, cyanidin, and pelargonidin-3-O-galactosides. On the contrary, consumption of whole blueberry powder and isolated anthocyanins from blueberry and strawberry yields a mixed result. In addition, high-fat diet mice fed with whole blueberry powder have increased body weight and adiposity relative to high-fat-fed controls [127]. Inversely, the study shows that the obese mice fed with isolated anthocyanins from the fruits reduced weight gain and body fat, but the differences were not always statistically significant. The authors also tested the purified anthocyanins and blueberry juice for the ability to prevent obesity by preparing a dose of 0.2 mg/ml anthocyanin in drinking water (0.49 mg/mouse/day). The finding shows that consumption of the purified anthocyanins suppressed the rate of fat deposition. Also, consumption of blueberry juice (2.8 ml/mouse/day; 5.3 mg of anthocyanin/mouse/day) was not as effective as the purified anthocyanins in preventing deposition of fat in the body. Moreover, lower serum leptin concentrations had been consistently observed in the purified blueberry anthocyanins (1.0 mg/ml) fed to obese mice for 72 days, which reduces the development of obesity [128].
Anthocyanin pigments are important nutraceuticals in maintaining good vision. Anthocyanin-rich berries are traditionally known for the goodness to eyes and are often associated with night vision. Most of the berries have high anthocyanins content. Oral administration of bilberry extract (contained about 39% anthocyanins) to six weeks old C57BL/6 mice has been shown to prevent impairment of photoreceptor cell function during retinal inflammation [91]. In another study, 132 patients with normal tension glaucoma were supplemented with two anthocyanins capsules (60.0 mg anthocyanin in each capsule) from bilberry daily and have improved visual function, based on the Humphrey visual field test and minimal angle of resolution best-corrected visual acuity assessment [90].
Some other berries demonstrate a protective effect for eyesight. Blackcurrant anthocyanin supplementation (50 mg/day) for 24 months increased ocular blood flow in 19 patients with open-angle glaucoma, however, there were no significant changes in the intraocular pressure [95]. Supplementation of anthocyanins (50 mg/kg body weight) from the seed coat of black soybean to N-methyl-N-nitrosourea-induced retinal degenerative rats also prevents retinal degeneration [94], and also suppresses human lens epithelial cell death under hydrogen peroxide-induced oxidative stress by 50–200 μg/ml of the extract [93]. Anthocyanin also predominates around 70% in purple corn seed [124], where purple corn seed extract decreases lens opacity together with lower malonaldehyde levels [92].
Anti-obesity effect
Anthocyanidin and anthocyanin pigments possess anti-obesity properties. Based on a previous study, obese mice fed a diet rich in cyanidin-3-glucoside from purple corn for 12 weeks have reduced body weight, as well as decreases in white and brown adipose tissue weights [100]. The study demonstrates that hyperglycemia, hyperinsulinemia, hyperleptinemia, and an increase in the tumor necrosis factor (TNF-a) mRNA level occurred in the obese rats are normalized when treated with purple corn diet. The purple corn also suppresses mRNA levels of enzymes involved in fatty acid and triacylglycerol synthesis and lowered sterol regulatory element binding protein-1 mRNA level in the white adipose tissue. These downregulations may contribute to a low triacylglycerol accumulation in white adipose tissue.
Obesity is strongly associated with adipocyte dysfunction. Therefore, regulation of protein secretion from adipocyte or the adipocyte-specific gene expression is one of the most important targets for prevention of obesity. Tsuda and his research team further investigated the potency of anthocyanins, particularly cyanidin and cyanidin-3-glucoside on isolated rat adipocytes for anti-obesity effect. He demonstrates that the adipocytes treated with anthocyanins have increased adiponectin and leptin secretions and upregulated adipocyte-specific gene expression without activation of PPARγ in the isolated rat adipocytes [99]. Gene expression of adiponectin is also upregulated in white adipose tissue of the anthocyanin-treated mice. The increased phosphorylation of AMPK may be associated with these changes, and the monophosphate/adenosine triphosphate ratio is significantly decreased by the administration of anthocyanins.
As previously reported by Tsuda et al., adipocyte gene expression is not thoroughly studied. A further examination of gene expression profile in isolated rat adipocytes treated with anthocyanins (100 nM cyanidin-3-glucoside or cyanidin) has been performed in vitro [125]. Within 24 h, a total of 633 genes and 427 genes were upregulated (1.5-fold) by the treatment of adipocytes with cyanidin-3-glucoside and cyanidin, respectively. The upregulated genes include lipid metabolism and signal transduction-related genes. However, the altered genes are partially different when comparing the cyanidin-3-glucoside and cyanidin treated groups. They also report that treatment of adipocytes with cyanidin-3-glucoside and cyanidin upregulated hormone sensitive lipase and enhanced lipolytic activity based on the microarray data. Even though the findings have identified new responsive genes with potentially important functions in adipocytes related to obesity, additional investigation is needed. In vivo adipocytes are not likely to be exposed to the anthocyanidin due to its instability in the culture.
Another study found that the ameliorated obese mice (C57BL/6) fed with Cornelian cherries (Cornus mas) containing anthocyanins (1 g/kg of high fat diet) for eight weeks had a 24% decrease in weight gain and decreased lipid accumulation in the liver, as well as a significant decrease in liver triacylglycerol concentration, independent of food intake [126]. The diet containing a mixture of delphinidin, cyanidin, and pelargonidin-3-O-galactosides. On the contrary, consumption of whole blueberry powder and isolated anthocyanins from blueberry and strawberry yields a mixed result. In addition, high-fat diet mice fed with whole blueberry powder have increased body weight and adiposity relative to high-fat-fed controls [127]. Inversely, the study shows that the obese mice fed with isolated anthocyanins from the fruits reduced weight gain and body fat, but the differences were not always statistically significant. The authors also tested the purified anthocyanins and blueberry juice for the ability to prevent obesity by preparing a dose of 0.2 mg/ml anthocyanin in drinking water (0.49 mg/mouse/day). The finding shows that consumption of the purified anthocyanins suppressed the rate of fat deposition. Also, consumption of blueberry juice (2.8 ml/mouse/day; 5.3 mg of anthocyanin/mouse/day) was not as effective as the purified anthocyanins in preventing deposition of fat in the body. Moreover, lower serum leptin concentrations had been consistently observed in the purified blueberry anthocyanins (1.0 mg/ml) fed to obese mice for 72 days, which reduces the development of obesity [128].
DonTesla
Well-Known Member
Antimicrobial
Polyphenolic compounds including anthocyanins possess antimicrobial activity against a wide range of microorganisms, especially in inhibiting the growth of food-borne pathogens [129]. Anthocyanins exhibit antimicrobial activity through several mechanisms, such as induced cell damage by destroying the cell wall, membrane, and intercellular matrix [101].
Based on a previous study, maqui berry extracts had antibacterial activity with the highest sensitivity to Aeromonas hydrophilia and Listeria innocua [102]. These bacteria are commonly associated with refrigerated foods as indicators of pathogenic microorganisms or as spoilage microorganisms [130]. Côté et al. [103] report that cranberry extract had antibacterial activity towards Enterococcus faecium resistant to vancomycin, Pseudomonas aeruginosa, Staphylococcus aureus, and Escherichia coli. The antibacterial activity of cranberry extract is not based on its low pH, but it is believed due to the other specific bioactive components, such as anthocyanins and flavonols in cranberry extracts after pH adjusted to 7.
Anthocyanin-rich extracts, such as blueberry, raspberry, blackcurrant, and strawberry extracts, inhibit Gram-negative bacteria but not Gram-positive bacteria [104]. This variation may be due to the different structures of cell wall between Gram-negative and Gram-positive bacteria, in which the outer membrane of Gram-negative bacteria acts as a preventive barrier against hydrophobic compounds but not on hydrophilic compounds [131]. These antimicrobial activities of anthocyanin-containing extracts are possibly due to the multiple mechanisms and synergistic effects of various phytochemicals in the extracts, including anthocyanins, weak organic acids, phenolic acids, and their mixtures of different chemical forms [132]. Thus, the antimicrobial effect of chemically complex compounds instead of solely anthocyanins should be extensively analyzed. Also, anthocyanins in purple, red, and blue-colored fruits and vegetables are the main bioactives in preventing microbial infection by several mechanisms.
Neuroprotective effect of anthocyanins
The term, ‘neuroprotection’, has been defined as the protection of nerve cells from oxidative injury and neurotoxicity, which interferes with the ischemic cascade. A neuroprotective agent is a drug or natural compound that prevents the nervous system from secondary injuries. The neuroprotective effects of anthocyanins have been evaluated based on the in vitro and in vivo studies. Most of the in vitro studies are performed by applying cell cultures, whereas in vivo studies are carried out based on animal models.
The neuroprotective findings from selected studies are reported in this review. An in vitro study [133] shows the neuroprotective effect of cyanidin-3-glucoside and its aglycone against hydrogen peroxide-induced oxidative stress in human neuronal cells (SH-SY5Y). The results demonstrate that SH-SY5Y cells pretreated with 100 µM cyanidin and cyanidin-3-glucoside significantly increased total antioxidant activity of membrane and cytosolic fraction from the cells; cyanidin also significantly increased the percentages of mitochondrial functioning and inhibited DNA fragmentation induced by hydrogen peroxide.
Based on a previous study, cyanidin-3-glucoside (2 mg/kg body weight) isolated from Prunus cerasus fruit inhibited apoptosis-inducing factor from mitochondria under oxidative stress but did not block the release of cytochrome c against permanent middle cerebral artery occlusion in the cortical neurons isolated from adult mice brain [134]. Besides the reported findings, the cyanidin-3-glucoside treated mice had brain superoxide level lower than the control (0.9% normal saline), as well as with better neurological test scores.
In another study, male Sprague-Dawley rats with traumatic spinal cord injury that received 400 mg/kg body weight of cyanidin-3-glucoside had a significantly improved blood-brain barrier score by 16.7%, platform hang by 40.0%, and hind foot bar grab by 30.8% compared to vehicle treated control, as well as significant reductions in superoxide level of the spinal cords and lesion volume in the lesion periphery, and a significant increased in motor neuron cell numbers of the anterior horn in lesion periphery [135]. The data from a mouse model of late pregnancy reveals that intraperitoneal injection of cyanidin-3-glucoside blocks ethanol-mediated glycogen synthase kinase-3β by inducing phosphorylation at serine 9 and reduces the phosphorylation at tyrosine 216 [136]. The compound also ameliorates ethanol-induced oxidative stress by inhibiting expression of malondialdehyde (MDA). The study concludes that cyanidin-3-glucoside prevents neurotoxicity of ethanol.
As reported in the literature, cyanidin-3-O-ß-d-glucopyranoside isolated from mulberry extract had a neuroprotective effect on PC12 pheochromocytoma cells through inhibiting cerebral ischemic damage induced by oxygen-glucose deprivation when the cells were exposed to hydrogen peroxide (150 µM) for 24 h [137]. The researchers also developed a mouse-brain-injury model of transient middle-cerebral artery occlusion, where the mice were supplemented with cyanidin-3-O-β-d-glucopyranoside and mulberry fruit extract. The result demonstrates a reduction in the infarct volume of the brain by 18% and 26%, respectively, and both supplemented groups had a lesser number of myeloperoxidase-positive cells than the ischemic control group in the striatum and cortex of the injured brain.
Based on the previous findings, most of the studies report the neuroprotective effects of cyanidin and its glycosides. Limited study has been done to determine the neuroprotective benefits of other anthocyanidins and anthocyanins. Kim et al. [138] show the neuroprotective effect of three major anthocyanins (a mixture of cyanidin-3-glucoside, delphinidin-3-glucoside, and petunidin-3-glucoside) isolated from black soybean, against hydrogen peroxide-induced cell death. They conclude that the human brain neuroblastoma SK-N-SH cells treated with the purified anthocyanin mixture (1–25 μg/ml) had a significant reduction in intracellular ROS level in a dose-dependent manner. The anthocyanins also inhibited ROS-dependent activation of apoptosis signal-regulating kinase 1 (ASK1)–JNK/p38 pathways, stimulated expression of heme oxygenase 1, and upregulated sialidase 1 (also known as Neu1) gene expression. Based on this evidence, anthocyanins obtained from plants have a neuroprotective effect.
Source:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5613902/
Polyphenolic compounds including anthocyanins possess antimicrobial activity against a wide range of microorganisms, especially in inhibiting the growth of food-borne pathogens [129]. Anthocyanins exhibit antimicrobial activity through several mechanisms, such as induced cell damage by destroying the cell wall, membrane, and intercellular matrix [101].
Based on a previous study, maqui berry extracts had antibacterial activity with the highest sensitivity to Aeromonas hydrophilia and Listeria innocua [102]. These bacteria are commonly associated with refrigerated foods as indicators of pathogenic microorganisms or as spoilage microorganisms [130]. Côté et al. [103] report that cranberry extract had antibacterial activity towards Enterococcus faecium resistant to vancomycin, Pseudomonas aeruginosa, Staphylococcus aureus, and Escherichia coli. The antibacterial activity of cranberry extract is not based on its low pH, but it is believed due to the other specific bioactive components, such as anthocyanins and flavonols in cranberry extracts after pH adjusted to 7.
Anthocyanin-rich extracts, such as blueberry, raspberry, blackcurrant, and strawberry extracts, inhibit Gram-negative bacteria but not Gram-positive bacteria [104]. This variation may be due to the different structures of cell wall between Gram-negative and Gram-positive bacteria, in which the outer membrane of Gram-negative bacteria acts as a preventive barrier against hydrophobic compounds but not on hydrophilic compounds [131]. These antimicrobial activities of anthocyanin-containing extracts are possibly due to the multiple mechanisms and synergistic effects of various phytochemicals in the extracts, including anthocyanins, weak organic acids, phenolic acids, and their mixtures of different chemical forms [132]. Thus, the antimicrobial effect of chemically complex compounds instead of solely anthocyanins should be extensively analyzed. Also, anthocyanins in purple, red, and blue-colored fruits and vegetables are the main bioactives in preventing microbial infection by several mechanisms.
Neuroprotective effect of anthocyanins
The term, ‘neuroprotection’, has been defined as the protection of nerve cells from oxidative injury and neurotoxicity, which interferes with the ischemic cascade. A neuroprotective agent is a drug or natural compound that prevents the nervous system from secondary injuries. The neuroprotective effects of anthocyanins have been evaluated based on the in vitro and in vivo studies. Most of the in vitro studies are performed by applying cell cultures, whereas in vivo studies are carried out based on animal models.
The neuroprotective findings from selected studies are reported in this review. An in vitro study [133] shows the neuroprotective effect of cyanidin-3-glucoside and its aglycone against hydrogen peroxide-induced oxidative stress in human neuronal cells (SH-SY5Y). The results demonstrate that SH-SY5Y cells pretreated with 100 µM cyanidin and cyanidin-3-glucoside significantly increased total antioxidant activity of membrane and cytosolic fraction from the cells; cyanidin also significantly increased the percentages of mitochondrial functioning and inhibited DNA fragmentation induced by hydrogen peroxide.
Based on a previous study, cyanidin-3-glucoside (2 mg/kg body weight) isolated from Prunus cerasus fruit inhibited apoptosis-inducing factor from mitochondria under oxidative stress but did not block the release of cytochrome c against permanent middle cerebral artery occlusion in the cortical neurons isolated from adult mice brain [134]. Besides the reported findings, the cyanidin-3-glucoside treated mice had brain superoxide level lower than the control (0.9% normal saline), as well as with better neurological test scores.
In another study, male Sprague-Dawley rats with traumatic spinal cord injury that received 400 mg/kg body weight of cyanidin-3-glucoside had a significantly improved blood-brain barrier score by 16.7%, platform hang by 40.0%, and hind foot bar grab by 30.8% compared to vehicle treated control, as well as significant reductions in superoxide level of the spinal cords and lesion volume in the lesion periphery, and a significant increased in motor neuron cell numbers of the anterior horn in lesion periphery [135]. The data from a mouse model of late pregnancy reveals that intraperitoneal injection of cyanidin-3-glucoside blocks ethanol-mediated glycogen synthase kinase-3β by inducing phosphorylation at serine 9 and reduces the phosphorylation at tyrosine 216 [136]. The compound also ameliorates ethanol-induced oxidative stress by inhibiting expression of malondialdehyde (MDA). The study concludes that cyanidin-3-glucoside prevents neurotoxicity of ethanol.
As reported in the literature, cyanidin-3-O-ß-d-glucopyranoside isolated from mulberry extract had a neuroprotective effect on PC12 pheochromocytoma cells through inhibiting cerebral ischemic damage induced by oxygen-glucose deprivation when the cells were exposed to hydrogen peroxide (150 µM) for 24 h [137]. The researchers also developed a mouse-brain-injury model of transient middle-cerebral artery occlusion, where the mice were supplemented with cyanidin-3-O-β-d-glucopyranoside and mulberry fruit extract. The result demonstrates a reduction in the infarct volume of the brain by 18% and 26%, respectively, and both supplemented groups had a lesser number of myeloperoxidase-positive cells than the ischemic control group in the striatum and cortex of the injured brain.
Based on the previous findings, most of the studies report the neuroprotective effects of cyanidin and its glycosides. Limited study has been done to determine the neuroprotective benefits of other anthocyanidins and anthocyanins. Kim et al. [138] show the neuroprotective effect of three major anthocyanins (a mixture of cyanidin-3-glucoside, delphinidin-3-glucoside, and petunidin-3-glucoside) isolated from black soybean, against hydrogen peroxide-induced cell death. They conclude that the human brain neuroblastoma SK-N-SH cells treated with the purified anthocyanin mixture (1–25 μg/ml) had a significant reduction in intracellular ROS level in a dose-dependent manner. The anthocyanins also inhibited ROS-dependent activation of apoptosis signal-regulating kinase 1 (ASK1)–JNK/p38 pathways, stimulated expression of heme oxygenase 1, and upregulated sialidase 1 (also known as Neu1) gene expression. Based on this evidence, anthocyanins obtained from plants have a neuroprotective effect.
Source:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5613902/
SSGrower
Well-Known Member
When going from cabinet to tent I decided to go with positive pressure ventilation to avoid collapsing the sides of the tent, since the fan operation is based on temperature or humidity it does not run all the time, only when either of those 2 parameters are out. The question is "how does (or does it?) this fluction actually impact any photochemical reactions going on?"View attachment 4165378
Note the expression here.. Various hues of pink, purple, magenta and black "Secretory cells"..
This time we had no negative pressure and no VP drops.. And as predicted ..no hypodermal cells affected.
Also no surprise that the basal cell (see below) also remained unaffected, as usual, thus far anyway
However, the interesting thing is that We were able to achieve this much expression with our water only soil recipe .. and the real kicker, no uv!!
.
View attachment 4165379
The pressure changes with the fan are significant enough to impact rh, and therfore vapor pressure and vpd. I garden at 7800 ft elevation so water holding capacity of air is less due to lower barrometric pressure (less mass of water for same rh reading) than at sea level. In my mind it seems that there should be some correlation to the vp of the terpenes or canibinoids and the barrometric pressure of the garden. If I could hold barrometric pressure constant I would have a baseline to go off, but alas, I am in a garage without that kind of budget.
Would be interisted to know what the atmospheric pressure is in your test garden.
DonTesla
Well-Known Member
Heres how it works i think..
Imo...Positive pressure.. including too high of humidity or too much co2 at night .. can have repressive effects .. See it as push-back if you will.. A positive, pressure and presence not conducive to flow of room-pull / plant-push..
Whereas negative pressure can have a drawing or pulling effect.. See film world war z when the grenade goes off in the plane.. If we want our plants to express fully and release the super freak traits (like the plane releasing zombies), its negative pressure that will give you pull. However, I think the pull is limited to hypodermal cells only and is blocked somehow by the basal cell.. Which I call the gateway cell to the crown cells.. The quarterback of the trichome..
I thought it was genetic predisposition with uv being thee major factor at play..however further trials were able to get entire rooms to express hypodermal pull / anthocyanin / pink or purple stalks, with various companies and genetic lines from various regions and eras, being tested at the same time.. Under various spectrums as well.. And all of them were able to get there..but the army of various strains could not be pushed beyond that.. no heads or secretary cells in other words would turn pink or express any anthocyanin for us ..
Ongoing Conclusion:
all cannabis is not only able to express anthocyanin, but should, and is meant to carry it, and when given the chance to, it will, since it wants to shine in its most abundant form... which can be aided and triggered especially with the presence of negative pressure, vp build ups/drops, living soil and imo, the biggie, full-spectrum compost ..
However, hypodermal expression is much easier to achieve than secretary cell expression, which may require genetic predisposition, as well as other triggers, or cheat codes which I am experimenting to discover, that will activate dormant DNA.
Imo...Positive pressure.. including too high of humidity or too much co2 at night .. can have repressive effects .. See it as push-back if you will.. A positive, pressure and presence not conducive to flow of room-pull / plant-push..
Whereas negative pressure can have a drawing or pulling effect.. See film world war z when the grenade goes off in the plane.. If we want our plants to express fully and release the super freak traits (like the plane releasing zombies), its negative pressure that will give you pull. However, I think the pull is limited to hypodermal cells only and is blocked somehow by the basal cell.. Which I call the gateway cell to the crown cells.. The quarterback of the trichome..
I thought it was genetic predisposition with uv being thee major factor at play..however further trials were able to get entire rooms to express hypodermal pull / anthocyanin / pink or purple stalks, with various companies and genetic lines from various regions and eras, being tested at the same time.. Under various spectrums as well.. And all of them were able to get there..but the army of various strains could not be pushed beyond that.. no heads or secretary cells in other words would turn pink or express any anthocyanin for us ..
Ongoing Conclusion:
all cannabis is not only able to express anthocyanin, but should, and is meant to carry it, and when given the chance to, it will, since it wants to shine in its most abundant form... which can be aided and triggered especially with the presence of negative pressure, vp build ups/drops, living soil and imo, the biggie, full-spectrum compost ..
However, hypodermal expression is much easier to achieve than secretary cell expression, which may require genetic predisposition, as well as other triggers, or cheat codes which I am experimenting to discover, that will activate dormant DNA.
SSGrower
Well-Known Member
Not seen the movie but I get what you are saying.
I have come to a similar conclusion on uv but will continue to experimint with it and red/far red/ir as in addition to full spectrum soil, I belive full spectrum light holds a key. I also saw some interisting results with the 10/10 light cycle and will continue experiminting with dli that way.
I have come to a similar conclusion on uv but will continue to experimint with it and red/far red/ir as in addition to full spectrum soil, I belive full spectrum light holds a key. I also saw some interisting results with the 10/10 light cycle and will continue experiminting with dli that way.
DonTesla
Well-Known Member
I agree that full spectrum lighting is a prerequisite to maximal glandular and compound expression, although I believe I have eliminated UV itself as the main factor..Not seen the movie but I get what you are saying.
I have come to a similar conclusion on uv but will continue to experimint with it and red/far red/ir as in addition to full spectrum soil, I belive full spectrum light holds a key. I also saw some interisting results with the 10/10 light cycle and will continue experiminting with dli that way.
it plays a support role, and is not the conductor per se.
SSGrower
Well-Known Member
Full contact gardening!
Must have been one of those shaped charge grenades to just poke a hole in the plan and not split it apart?
I will have to do some dinking with an altimeter to see to what degree the positive pressure ventilation raises the pressure, but, honestly this last run was probably my best since "back in the day".
Let me know if you need any help balancing the air locks for your greenhouses. It's part of what I used to do for the potent compound labs and clean rooms BITD.
DonTesla
Well-Known Member
Well, its still a movie after all, flaws with every film, but I think it illustrates the power of negative pressure nicely and the pulling effect.Full contact gardening!
Must have been one of those shaped charge grenades to just poke a hole in the plan and not split it apart?
I will have to do some dinking with an altimeter to see to what degree the positive pressure ventilation raises the pressure, but, honestly this last run was probably my best since "back in the day".
Let me know if you need any help balancing the air locks for your greenhouses. It's part of what I used to do for the potent compound labs and clean rooms BITD.
Having your best round in a long time though, thats huge.. always great when your round goes well.
And cool, very good to know!